ENZIM
Karbohidrat yaitu senyawa organik terdiri dari unsur karbon, hidrogen, dan oksigen. Terdiri atas unsur C, H, O dengan perbandingan 1 atom C, 2 atom H, 1 atom O. karbohidrat banyak terdapat pada tumbuhan dan binatang yang berperan struktural & metabolik. sedangkan pada tumbuhan untuk sintesis CO2 + H2O yang akan menghasilkan amilum/selulosa, melalui proses fotosintesis, sedangkan binatang tidak dapat menghasilkan karbohidrat sehingga tergantung tumbuhan. Karbohidrat merupakan sumber energi dan cadangan energi, yang melalui proses metabolisme. Banyak sekali makanan yang kita makan sehari hari adalah sumber karbohidrat seperti : nasi/beras, singkong, umbi-umbian, gandum, sagu, jagung, kentang, dan beberapa buah-buahan lainnya, dll. Di dalam tubuh karbohidrat dapat dibentuk dari beberapa asam amino dan sebagian dari gliserol lemak. Sumber karbohidrat nabati dalam glikogen bentuk glikogen, hanya dijumpai pada otot dan hati dan karbohidrat dalam bentuk laktosa hanya dijumpai di dalam susu.
Metabolisme merupakan total reaksi kimia yang terjadi didalam tubuh makhluk hidup untuk kelangsungan kehidupannya. Secara keseluruhan reaksi-reaksi tersebut bertanggungjawab untuk menjaga availabilitas organisme. Reaksi-reaksi tersebut secara sendiri-sendiri mungkin tidak penting, tetapi secara keseluruhan dalam jejaring akan membentuk rangkaian yang sangat dibutuhkan untuk keseimbangan fungsi biokimia. Adanya gangguan pada salah satu reaksi akan menyebabkan abnormalitas metabolisme.Reaksi-reaksi metabolisme dapat dibagi menjadi dua sesuai dengan tujuan reaksinya, yaitu katabolisme dan anabolisme. Katabolisme merupakan reaksi peluruhan (degradasi) yang menghasilkan energi, sedang anabolisme merupakan reaksi sintesis yang memerlukan energi. Keduanya berjalan secara seimbang sesuai dengan fungsi dan kebutuhan hidup organisme.
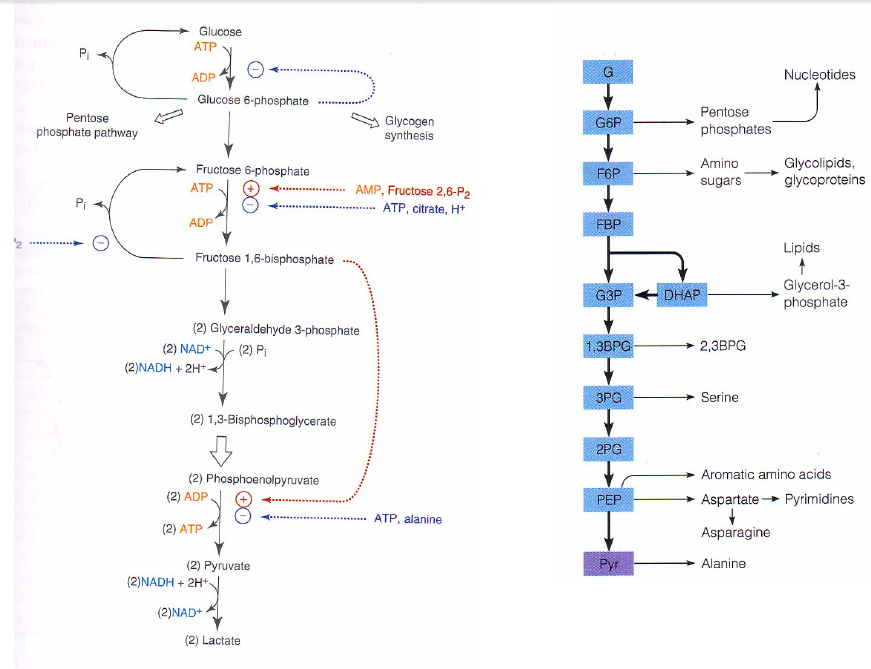
Glukosa merupada senyawa golongan karbohidrat yang merupakan sumber energi utama bagi makhluk hidup karena glukosa berasal dari proses fotosintesis yang mengkonversi energi matahari menjadi energi kimia. Energi yang terkandung dalam senyawa glukosa selanjutnya akan ditransformasi melalui serangkaian reaksi katabolisme yang dinamakan glikolisis. Glikolisis terjadi di dalam sitosol di dalam sel yang menghasilkan senyawa luruhan dan energi konversi dalam bentuk senyawa kimia yang lain (ATP).
Glikolisis adalah proses pemecahan glukosa pada tingkat sel. Pada makalah ini saya menjelaskan tahap-tahap glikolisis yang detail setiap tahap dalam proses biokimia yang merupakan bagian dari respirasi selular. Akan melalui sepuluh tahap akan memberi Anda wawasan tentang bagaimana reaksi biokimia yang kompleks dan terkoordinasi dengan baik.
Glikolisis adalah rincian sistematis glukosa dan gula lain untuk kekuatan proses respirasi selular. Ini adalah reaksi biokimia universal yang terjadi dalam setiap organisme uniseluler atau multiseluler yang hidup respires aerobik dan anaerobik. Ada jalur metabolik di mana proses ini terjadi. Tahap glikolisis pada jalur tertentu yang disebut embden-Meyerhof-Parnus jalur. Proses ini adalah bagian kecil dari siklus respirasi seluler dan metabolisme tubuh secara keseluruhan, diarahkan untuk menciptakan ATP (Adenosine Triphosphate) yang merupakan mata uang energi tubuh.
Glikolisis adalah pemecahan glukosa menjadi asam piruvat atau asam laktat. Jalur ini terutama terjadi dalam otot bergaris, yang dimaksudkan untuk menghasilkan energi (ATP). Apabila glikolisis terjadi dalam suasana anaerobik maka akan berakhir dengan asam laktat, dan mengha-silkan dua ATP, apabila dalam keadaan aerobik berakhir menjadi asam piruvat dengan 8 ATP.
Berikut Penjelasan Singkat langkah-langkah dalam metabolisme karbohidrat :
GLIKOLISIS yaitu: dimana glukosa dimetabolisme menjadi piruvat (aerob) menghasilkan energi (8 ATP)atau laktat (anerob)menghasilkan (2 ATP).
GLIKOGENESIS yaitu: proses perubahan glukosa menjadi glikogen. Di Hepar/hati berfungsi: untuk mempertahankan kadar gula darah. sedangkan di Otot bertujuan: kepentingan otot sendiri dalam membutuhkan energi.
GLIKOGENOLISIS yaitu : proses perubahan glikogen menjadi glukosa. atau kebalikan dari GLIKOGENESIS.
JALUR PENTOSA FOSFAT yaitu : hasil ribosa untuk sintesis nukleotida, asam nukleat dan equivalent pereduksi (NADPH) (biosintesis asam lemak dan lainnya.)
GLUKONEOGENESIS : senyawa non-karbohidrat (piruvat, asam laktat, gliserol, asam amino glukogenik) menjadi glukosa.
Pengantar Glucose-6-phosphatase
Glucose 6-phosphatase (EC 3.1.3.9, G6Pase) merupakan enzim hirolisasi menjadi glucose-6-phosphate, menghasilkan reaksi grup phospate dan bebas glukosa. Glukosa yang diambil dari sel melalui transport glukosa membran protein. Katalisator sempurna lengkap terakhir menjadi glukoneogenesis dan glikogenolisis yang memainkan peran dalam regulasi homeostatik dalam tingkat glukosa darah. G6Pase lengkap dengan banyak komponen protein, termasuk transpor untuk G6P,glukosa,dan phosphate. Fungsi utama phosphate yang ditunjukkan oleh G6Pase katalisator subunit. Pada manusia, tiga (3) isozymem katalisator unit yaitu: G6Pase-α, dikode oleh G6PC; IGRP, dikode oleh G6PC2; dan G6Pase-β, dikode oleh G6PC3. G6Pase-α dan G6Pase-β kedua-duanya merupakan fungsi fospohidrolasi, dan kemiripan aktif tempat struktur,topologi, mekanisme aktif dan peralatan kinetik dengan menjadi hidrolisis G6P. Memperbandingkan,IGRP hampir tidak ada aktifitas hidrolisis, dan memainkan perbedaan mendorong sekresi pankreas insulin.

Secara rinci, tahap-tahap dalam lintasan glikolisis adalah sebagai berikut (pada setiap tahap, lihat dan hubungkan dengan Gambar Lintasan detail metabolisme karbohidrat):
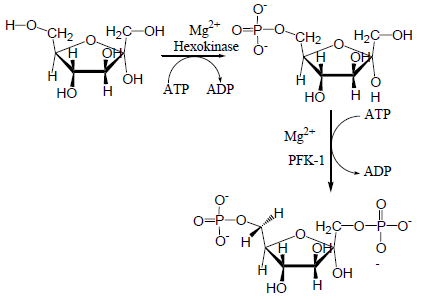
1. Glukosa masuk lintasan glikolisis melalui fosforilasi menjadi glukosa-6 fosfat dengan dikatalisir oleh enzim heksokinase atau glukokinase pada sel parenkim hati dan sel Pulau Langerhans pancreas. Proses ini memerlukan ATP sebagai donor fosfat. ATP bereaksi sebagai kompleks Mg-ATP. Terminal fosfat berenergi tinggi pada ATP digunakan, sehingga hasilnya adalah ADP. (-1P) Reaksi ini disertai kehilangan energi bebas dalam jumlah besar berupa kalor, sehingga dalam kondisi fisiologis dianggap irrevesibel. Heksokinase dihambat secara alosterik oleh produk reaksi glukosa 6-fosfat.
2. Glukosa 6-fosfat diubah menjadi Fruktosa 6-fosfat dengan bantuan enzim
fosfoheksosa isomerase dalam suatu reaksi isomerasi aldosa-ketosa. Enzim ini hanya bekerja pada anomer µ-glukosa 6-fosfat.
Glikolisis diawali dengan reaksi pembentukan senyawa glukosa 6-fosfat dari glukosa. Reaksi tersebut merupakan reaksi yang membutuhkan energi yang diambil dari pemutusan ikatan fosfat dari ATP. Reaksi ini dikatalisis oleh enzim heksokinase atau glukokinase.

Heksokinase dapat ditemukan dalam semua sel organisme. Enzim ini memiliki spesifitas katalitik yang rendah. Hampir semua monosakarida dapat difosforilasi. Aktivitasnya dapat dihambat oleh produknya, yaitu glukosa-6-fosfat.Glukokinase diitemukan di lever, memiliki spesifitas katalitik yang tinggi dan tidak dapat dihambat oleh glukosa-6-fosfat.. Enzim ini aktif bila kadar glukosa tinggi di dalam darah. Heksokinase memerlukan logam dengan ion divalent seperti Mg2+ untuk dapat teraktivasi. Ion logam ini dapat membentuk kompleks dengan ATP. Heksokinase diinhibisi oleh produk yang dikatalisis yaitu glukosa-6-phosphate. Konsentrasi yang tinggi dari glukosa-6-phosphate dapat memberikan signal bahwa sel tidak memerlukan lagi glukosa untuk dijadikan energi.
Isomerisasi glukosa 6-fosfat. Reaksi yang kedua adalah pembentukan isomer fruktosa 6-fosfat dari glukosa 6-fosfat. Reaksi ini dikatalisis oleh fosfoglukoisomerase.

Glukokinase terdapat dalam hepar dan pankreas. Mempunyai Km untuk D-glukosa jauh lebih tinggi dari enzim hexokinase. Glukokinase memerlukan glukosa lebih tinggi untuk menjadi aktif bila dibandingkan dengan heksokinase.
Berbeda dengan heksokinase glukokinase tidak dihambat oleh hasil reaksinya yaitu glukosa 6-P. Glukokinase berperan biasanya pada waktu kadar glukosa darah tinggi (sesudah makan). Pada penderita Diabetes Mellitus enzim ini jumlahnya berkurang.
Reaksi fosforilasi ini boleh dikatakan reaksi satu arah. Selanjutnya glukosa 6-P diubah menjadi fruktosa 6-P. Reaksi ini dikatalisis enzim fosfoheksosa isomerase, dimana terjadi aldosa-ketosa isomerasi. Hanya D-anomer dari glukosa 6-P yang bisa dipakai sebagai substrat. Reaksi ini merupakan reaksi bolak-balik.
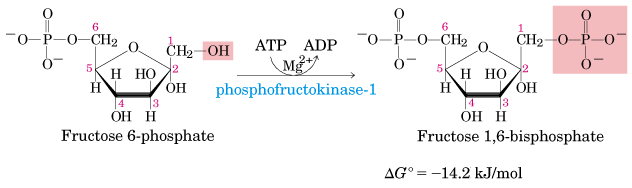
Reaksi selanjutnya adalah pembentukan fruktosa 1,6-difosfat oleh enzim fosfofruktokinase-1 (PFK-1). Reaksi ini boleh dikatakan reaksi satu arah. Enzim fosfofruktokinase-1 merupakan enzim yang bisa diinduksi. Enzim ini memegang peran yang penting dalam mengatur kecepatan glikolisis.
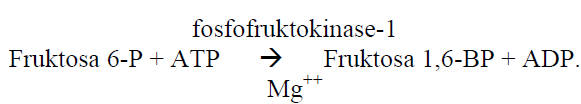
Aktifitas enzim ini meningkat apabila konsentrasi ADP, AMP, fosfat inorganik ( Pi ) meningkat. Enzim fosfofruktokinase-1 dihambat oleh ATP, asam sitrat dan 2,3-DP gliserat (dalam sel darah merah). Apabila pemakaian ATP meningkat (kadar ATP menurun) maka aktifitasnya meningkat, sebaliknya apabila kadar ATP tinggi aktifitas enzim tersebut menurun. Enzim ini juga dihambat oleh meningkatnya kadar asam lemak bebas, sehingga apabila senyawa ini meningkat dalam darah, yang akhirnya masuk ke dalam sel , maka pemakaian glukosa akan berkurang. Keadaan ini bisa terjadi pada waktu kelaparan, yang mana juga dapat terbentuk senyawa keton.
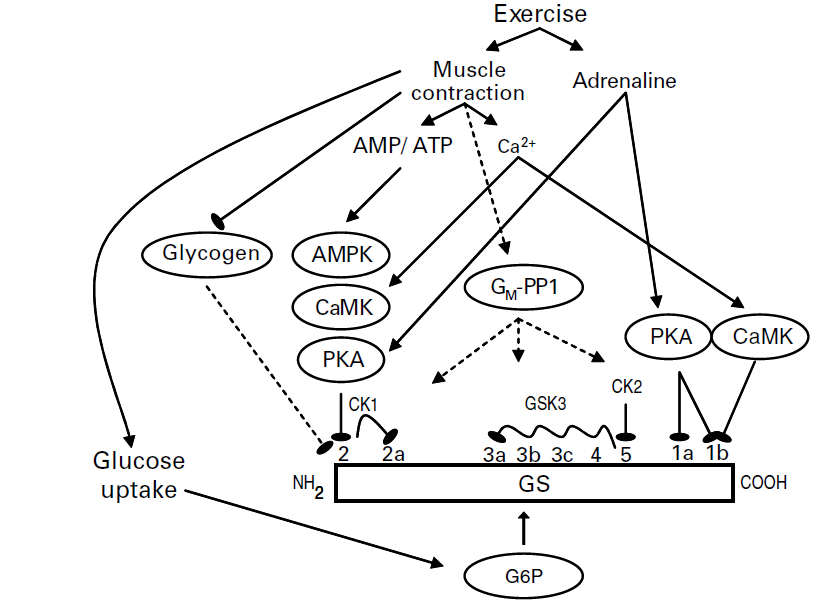
Gambar. 1. Peraturan rangka glikogen otot sintase (GS) aktivitas dan fosforilasi oleh latihan. Latihan memunculkan peningkatan kadar adrenalin darah dan kontraksi otot menyebabkan perubahan nukleotida dan ion kalsium tingkat intraselular. Adrenalin mengaktifkan cAMP-dependent protein kinase (AMPK) / protein kinase A (PKA), yang menurunkan aktivitas GS dengan langsung fosforilasi situs 2 dan secara tidak langsung fosforilasi perkemahan 3 dengan mekanisme yang belum diketahui (tidak ditunjukkan). The glycogenassociated protein fosfatase 1 (PP1) subunit GM, telah terbukti menjadi penting dalam kontraksi-aktivasi GS, tetapi mekanisme yang GM diaktifkan (jika sama sekali) oleh kontraksi otot belum teridentifikasi. Selain itu, tidak diketahui apakah PP1 istimewa dephosphorylates situs tertentu di GS. Glikogen menghambat aktivitas GS dengan menyebabkan fosforilasi situs 2 di GS, tapi mekanisme tetap sulit dipahami. Selanjutnya, meningkat konsentrasi ion kalsium dalam menanggapi kontraksi otot dan faktor ini bisa menyebabkan peningkatan perkemahan 2 fosforilasi GS dimediasi oleh ion kalsium / tergantung kalmodulin kinase (CaMK) atau dependent kinase-ion kalsium lainnya. Glukosa-6-fosfat (G6P) tingkat sering meningkat selama latihan sebagai akibat dari peningkatan ambilan glukosa dan peningkatan pemecahan glikogen. Perubahan tingkat allosterically mengaktifkan GS. Mengaktifkan dan faktor masing-masing menghambat. Efek yang tidak sepenuhnya dipahami. Enzim GS ditampilkan dalam bentuk diagram dengan indikasi situs fosforilasi yang berbeda dan kinase utama berpikir untuk bertindak di situs tersebut. Fosforilasi hirarki yang ditunjukkan pada situs 2a dan situs 3-4. Situs 2, 2a, 3a dan 3b adalah situs dianggap paling penting dalam regulasi aktivitas GS. GSK, GS kinase; CK1 dan 2, kinase kasein 1 dan 2. Untuk detail lebih lanjut dan referensi, lihat hal. 233.
Glukosa-6-fosfat Seperti disebutkan sebelumnya, G6P adalah stimulator langsung dan tidak langsung dari aktivitas GS. Meskipun efek alosterik langsung G6P in vivo tidak terdeteksi ketika aktivitas GS diukur in vitro, harus dipertimbangkan bahwa G6P mungkin memainkan peran penting dalam mengatur aktivitas GS selama latihan. Penyerapan glukosa otot meningkat dengan olahraga dan peningkatan ini sering menyebabkan peningkatan konsentrasi tingkat G6P. Dengan demikian, aktivasi alosterik dari GS selama latihan bisa sangat penting, seperti yang disarankan oleh Bloch et al. (1994). Kegiatan GS juga meningkat pada kontraksi otot rangka yang telah dirangsang untuk kontrak in vitro (diinkubasi) atau in situ (perfusi) tanpa adanya glukosa, menjaga kadar G6P sangat rendah atau bahkan tidak berubah (W Derave, komunikasi pribadi). Temuan ini menunjukkan bahwa peningkatan aktivitas GS diinduksi oleh latihan tidak tergantung pada peningkatan kadar G6P otot rangka, yang juga didukung oleh pengamatan pada kultur sel otot manusia (Montell et al. 1999)
Pengantar Fructose-1,6-bisphosphatase
Fructose-1,6-bisphosphatase (FBPase; EC 3.1.3.11), dimana hidrolisis dikatalisis oleh D-fructose-1,6-bisphosphate (FBP) menjadi D-fructose-6-phosphate (F6P) dan inorganic phosphate (Pi),yang merupakan kunci enzim glukogeogenesis. Seiring dengan fosfofruktokinase (EC 2.7.1.11), yang mengkatalisis reaksi sebaliknya, fosforilasi F6P selama glikolisis, searah FBPase mengatur fluks metabolisme gula. Enzim ini hanya mengkatalisis reaksi dalam satu arah masing-masing, dan diatur oleh metabolit seperti fruktosa 2,6-bifosfat sehingga aktivitas tinggi salah satu dari dua enzim disertai dengan rendahnya aktivitas yang lain. Lebih khusus, fruktosa 2,6-bifosfat allosterically menghambat fruktosa 1,6-bisphosphatase, tetapi mengaktifkan fosfofruktokinase-I. Fruktosa 1,6-bisphosphatase terlibat dalam banyak jalur metabolisme yang berbeda dan ditemukan di sebagian besar organisme. FBPase memerlukan ion logam untuk katalisis (Mg2 + dan Mn2 + yang disukai) dan enzim poten dihambat oleh Li +.
Reaksi selanjutnya adalah pembentukan fruktosa 1,6-difosfat oleh enzim fosfofruktokinase-1 (PFK-1). Reaksi ini boleh dikatakan reaksi satu arah. Enzim fosfofruktokinase-1 merupakan enzim yang bisa diinduksi. Enzim ini memegang peran yang penting dalam mengatur kece-patan glikolisis.

Aktifitas enzim ini meningkat apabila konsentrasi ADP, AMP, fosfat inorganik ( Pi ) meningkat. Enzim fosfofruktokinase-1 dihambat oleh ATP, asam sitrat dan 2,3-DP gliserat (dalam sel darah merah). Apabila pemakaian ATP meningkat (kadar ATP menurun) maka aktifitasnya meningkat, sebaliknya apabila kadar ATP tinggi aktifitas enzim tersebut menurun. Enzim ini juga dihambat oleh meningkatnya kadar asam lemak bebas, sehingga apabila senyawa ini meningkat dalam darah, yang akhirnya masuk ke dalam sel , maka pemakaian glukosa akan berkurang. Keadaan ini bisa terjadi pada waktu kelaparan, yang mana juga dapat terbentuk senyawa keton.
Peran fruktosa 2,6 bisfosfat (dalam hepar). Dalam hepar fruktosa 2,6-bisfosfat merupakan allosterik efektor positif yang paling kuat bagi enzim fosfofruktokinase1, dan merupakan inhibitor bagi enzim fruktosa 1,6-bisfosfatase ("enzim glukoneogenesis"). Fruktosa 2,6-bisfosfat menghilangkan pengaruh hambatan (inhibisi) ATP terhadap fosfofruktokinase-1, dan meningkatkan affinitas enzim ini terhadap fruktosa 6-P. Fruktosa 2,6-bisfosfat menghambat fruktosa-1,6-bisfosfatase dengan jalan meningkatkan harga Km untuk fruktosa-1,6-bisfosfat. Kadar fruktosa 2,6-bisfosfat dibawah pengaruh kontrol substrat dan hormonal.
Fruktosa 2,6-bisfosfat dibentuk dengan fosforilasi fruktosa 6-fosfat (fruktosa 6-P) yang dikatalisis enzim fosfofruktokinase-2 (PFK-2). Enzim yang sama bertanggung jawab juga terhadap pemecahan fruktosa 2,6 bisfosfat (F 2,6-BP), karena enzim ini mempunyai aktifitas fruktosa 2,6 bisfosfatase, namun enzim ini telah mengalami fosforilase menjadi PFK-2P (fosfo frukto kinase fosfat). Aktifitas bifungsi enzim fosfofruktokinase-2 ini dibawah pengaruh (kontrol) allosterik fruktosa 6-P. Apabila kadar senyawa ini meningkat, sebagai akibat meningkatnya kadar glukosa, maka akan meningkatkan aktifitas fosfofruktokinase-2 pada waktu yang sama aktifitas fosfatasenya menurun. Apabila kita memerlukan glukosa (pada waktu puasa), dengan perkataan lain glukosa darah menurun, glukagon akan menyebabkan terbentuknya cAMP, selanjutnya "cAMP- dependent protein kinase" teraktifasi, yang menyebabkan fosfofruktokinase-2 dihambat, sedangkan aktifitas fruktosa 2,6BPase meningkat, karena diubah menjadi PFK-2P.

Dalam keadaan dimana kadar glukosa meningkat, menyebabkan fruktosa 6P meningkat, ini memacu PFK-2 membentuk fruktosa 2,6 bisfosfat. Selanjutnya F 2,6 BP akan merangsang akti-fitas fosfofruktokinase-1 dan menghambat fruktosa 1,6 bisfosfatase.
Glukagon dan glukosa adalah dua efektor utama yang masing-masing, merangsang dan menghambat glukoneogenesis di hati dan memiliki efek sebaliknya pada glikolisis. Fruktosa 2,6-bifosfat dan AMP memiliki kesamaan sifat menghambat fruktosa-1,6-bisphosphatase dan merangsang fosfofruktokinase. Perubahan konsentrasi efektor ini karena itu bisa menjelaskan efek glukagon dan glukosa. Dengan demikian, konsentrasi fruktosa 2,6-bifosfat dikurangi secara bermakna dengan glukagon dan meningkat glukosa (1). Sebaliknya, konsentrasi AMP dalam hati meningkat dengan glukagon dan tidak terpengaruh oleh glukosa (9). Oleh karena itu, seseorang mencapai kesimpulan bahwa fruktosa 2,6-bifosfat
daripada AMP adalah utusan dari kedua glukosa dan glukagon dalam aksi mereka pada glukoneogenesis dan glikolisis. Penghambatan offructose-1, 6-bisphosphatase oleh fruktosa 2,6- bifosfat, bersama-sama dengan stimulasi fosfofruktokinase oleh efektor yang sama, menjelaskan juga peningkatan yang luar biasa dalam konsentrasi fruktosa 1,6-bifosfat dalam hati pada kondisi di mana fruktosa 2,6-bifosfat meningkat (1). Selanjutnya, bentuk sigmoidal substrat kurva kejenuhan fruktosa-1,6-bisphosphatase menunjukkan bahwa sekarang ada konsentrasi ambang fruktosa 1,6-bifosfat di mana ada sedikit konversi ke fruktosa 6-fosfat. Kesimpulan bahwa aktivitas fruktosa-1,6-bisphosphatase tidak diatur hanya dengan konsentrasi yang substrat telah dicapai sebelumnya (9). Ini juga telah dikenal selama bertahun-tahun (9-12) bahwa konsentrasi fruktosa 1,6-bifosfat di hati berkurang oleh administrasi glukagon. Hasil kami menunjukkan bahwa efek glukagon ini dimediasi oleh hilangnya fruktosa 2,6-bifosfat.
Peningkatan tingkat glikolisis yang terjadi pada glukosa yang tinggi konsentrasi menyiratkan tidak hanya fosfofruktokinase yang dirangsang tetapi juga bahwa aktivitas kinase piruvat meningkat secara paralel. Yang terakhir enzim sangat dirangsang oleh fruktosa 1,6-bifosfat. Meningkat offructose Konsentrasi 1,6-bifosfat yang merupakan hasil dari penghambatan fosfatase mungkin sinyal yang mengaktifkan piruvat kinase dan memungkinkan tingkat yang cepat konversi metabolit melalui jalur glikolitik. Catatan Ditambahkan pada Bukti. Kami baru-baru ini mengamati fruktosa bahwa 2,6-bifosfat juga merupakan inhibitor poten dari fruktosa-1,6-bisphosphatase dimurnikan dari ragi (Sigma) atau diekstrak dari daun bayam.
Metabolism of Hexoses other than glucose.
Fruktosa.
Sukrosa merupakan sumber bahan bakar utama makanan kita. Sukrosa merupakan disakarida dari fruktosa dan glukosa. Ada dua jalur untuk metabolisme fruktosa.
Yang pertama dan paling sederhana untuk jalur fruktosa katabolisme terjadi pada sel-sel otot di mana fruktosa adalah substrat untuk heksokinase yang menghasilkan fruktosa-6-fosfat, substrat untuk PFK-1. Hati, bagaimanapun, mengandung sedikit heksokinase. Sebaliknya mengandung glukokinase yang secara khusus phosphorylates glukosa. Hati memetabolisme fruktosa melalui jalur yang melibatkan enam enzim.
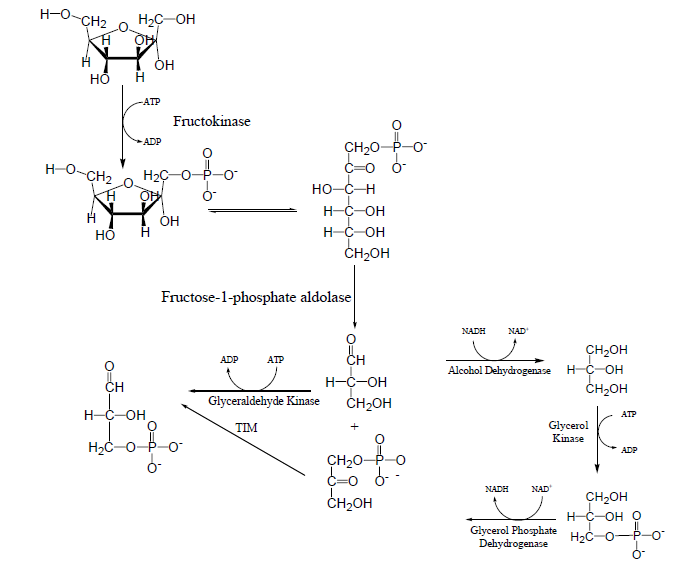
Langkah 1:
Dalam hati langkah pertama dari fruktosa katabolisme adalah fosforilasi fruktosa oleh fructokinase untuk membentuk fruktosa-1-fosfat. Perhatikan bahwa baik heksokinase atau fosfofruktokinase dapat memfosforilasi fruktosa-1-fosfat menjadi fruktosa-1,6-bifosfat.
Langkah 2:
Dalam hati kita menemukan kelas I aldolase yang merupakan isozim fruktosa bifosfat aldolase (Tipe aldolase A). Tipe A aldolase spesifik untuk substrat fruktosa-1,6-bifosfat. Isozim dari aldolase ditemukan di hati disebut aldolase Tipe B. Hal ini dapat memanfaatkan fruktosa-1-fosfat serta fruktosa-1,6-bifosfat sebagai substrat. Jadi aldolase Tipe B ditemukan dalam hati mengkatalisis pembelahan hidrolitik dari fruktosa-1-fosfat menjadi gliseraldehida dan dihidroksiaseton fosfat.
Langkah 3:
Dihidroksiaseton fosfat yang dihasilkan dapat dikonversi menjadi gliseraldehida 3-fosfat dengan reaksi dikatalisis oleh isomerase fosfat triose. Produk lain dari belahan dada hidrolitik, gliseraldehida dapat langsung terfosforilasi oleh kinase gliseraldehida untuk membentuk glyceraldehydes 3- fosfat.
Langkah 4: Ada jalur alternatif di mana glyceraldehydes dikurangi dengan alkohol dehidrogenase menjadi gliserol. Gliserol kemudian terfosforilasi oleh gliserol kinase untuk membentuk gliserol-3-fosfat. Gliserol 3-fosfat kemudian dioksidasi oleh gliserol fosfat dehidrogenase menjadi dihidroksiaseton fosfat yang tentu saja diubah menjadi glyceraldehydes 3-fosfat oleh isomerase fosfat triose.
Tahapan glikolisis yang akan menjelaskan reaksi fruktosa-1,6-bisfosfat oleh enzim fosfofruktokinase:
1. Glikolisis diawali dengan reaksi pembentukan senyawa glukosa 6-fosfat dari glukosa. Reaksi tersebut merupakan reaksi yang membutuhkan energi yang diambil
dari pemutusan ikatan fosfat dari ATP. Reaksi ini dikatalisis oleh enzim heksokinase atau glukokinase. Heksokinase dapat ditemukan dalam semua sel organisme. Enzim ini memiliki spesifitas katalitik yang rendah. Hampir semua monosakarida dapat difosforilasi. Aktivitasnya dapat dihambat oleh produknya, yaitu glukosa-6-fosfat. Glukokinase diitemukan di lever, memiliki spesifitas katalitik yang tinggi dan tidak dapat dihambat oleh glukosa-6-fosfat.. Enzim ini aktif bila kadar glukosa tinggi di dalam darah.
2. Isomerisasi glukosa 6-fosfat. Reaksi yang kedua adalah pembentukan isomer fruktosa 6-fosfat dari glukosa 6-fosfat. Reaksi ini dikatalisis oleh fosfoglukoisomerase.
3. Fosforilasi kedua. Reaksi fosforilasi fruktosa-6-fosfat menjadi fruktosa-1,6-bisfosfat oleh enzim fosfofruktokinase. Reaksi ini berjalan spontan dan merupakan rate limiting step pada proses glikolisis. Pada reaksi ini dibutuhkan 1 mol ATP dan
diregulasi secara ketat. Fosfofruktokinase dapat dihambat oleh ATP.
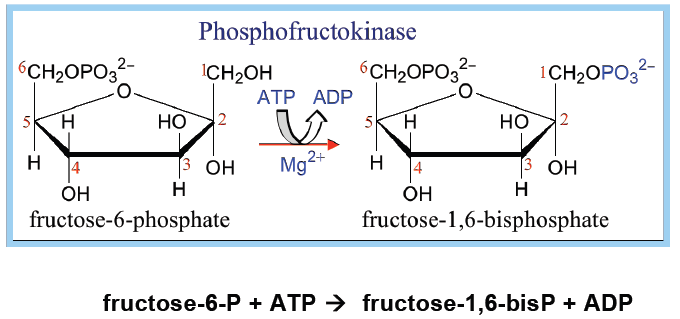
Fruktosa 6-fosfat diubah menjadi Fruktosa 1,6-bifosfat dengan bantuan enzim fosfofruktokinase. Fosfofruktokinase merupakan enzim yang bersifat alosterik sekaligus bisa diinduksi, sehingga berperan penting dalam laju glikolisis. Dalam kondisi fisiologis tahap ini bisa dianggap irreversible. Reaksi ini memerlukan ATP sebagai donor fosfat, sehingga hasilnya adalah ADP.(-1P)
Fruktosa 1,6-bifosfat dipecah menjadi 2 senyawa triosa fosfat yaitu gliserahdehid 3-fosfat dan dihidroksi aseton fosfat. Reaksi ini dikatalisir oleh enzim aldolase (fruktosa1,6-bifosfat aldolase).
Fructose-1,6-bisphosphatase berfungsi mengkatalisis reaksi pengubahan fruktosa-1,6-bifosfat menjadi fruktosa-6-posfat:
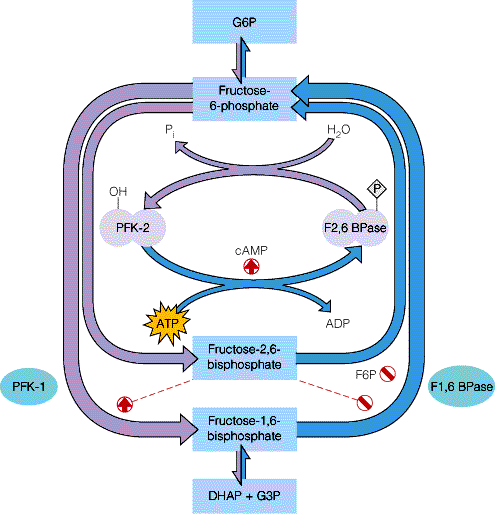
Inhibisi oleh fruktosa-2,6-bisfosfat
Fruktosa-2,6-bisfosfat merupakan inhibitor kompetitif. Proses inhibisi dapat terjadi karena fruktosa-1,6-bisfosfatase bersifat alosterik. Aktifitas enzim pada berbagai konsentrasi substrat dapat dilihat pada grafik di bawah ini :

Dari grafik tersebut diketahui bahwa pengaruh inhibisi Fruktosa-2,6-bisfosfat akan maksimum pada saat konsentrasi substrat kecil. Hal ini membuktikan bahwa Fruktosa-2,6-bisfosfat merupakan inhibitor kompetitif.
Inhibisi oleh AMP
Konsentrasi AMP dan ADP yang tinggi merupakan sinyal bahwa energi sel rendah. ADP menginhibisi piruvat karboksilase dan fosfoenolpiruvat karboksilase sementara AMP menginhibisi fruktosa 1,6 bisfosfatase. AMP mengaktivasi glikolisis melalui fosforfruktokinase. Dengan kata lain, ketika tingkat energi rendah, glukoneogenesis diinhibisi sementara glikolisis diaktivasi.
Inhibisi fructose-1,6-bisphosphatase oleh AMP bersifat unkompetitif terhadap fructose 1,6-bisphosphate pada keadaan tidak tersedia fructose 2,6-bisphosphate. Akan tetapi, bersifat nonkompetitif apabila fructose 2,6-bisphosphate tersedia. AMP tidak dapat berikatan dengan enzim kecuali dengan adanya salah satu dari senyawa fruktosa bisfosfat.
Inhibisi PFK oleh ATP
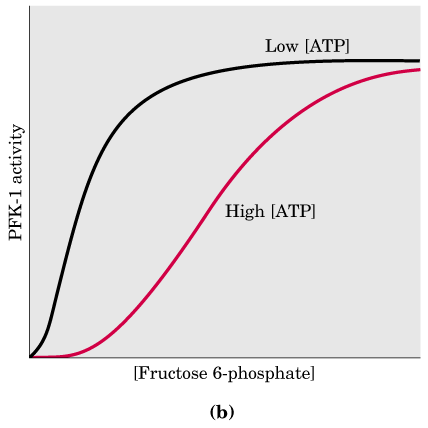
Pada diagram ini dapat ditunjukkan bahwa kerja enzim mencapai kerja maksimum pada pada saat keberadaan ATP rendah.
Ketika jumlah ATP dalam sel tinggi, sel tidak lagi membutuhkan produksi energi metabolik agar reaksi ini terjadi. Dalam hal ini, aktivitas PFK diinhibisi secara regulasi alosterik oleh ATP itu sendiri, menutup jalur katabolisme karbohidrat melalui jalur glikolisis ini.
Aktivasi PFK oleh ADP dan AMP
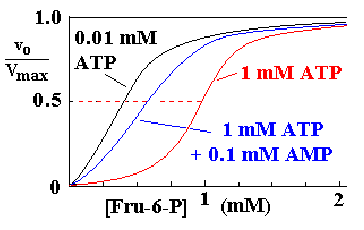
AMP dapat membalikan efek inhibitor dari ATP. Adanya AMP mengindikasikan bahwa energi dari sel sangat rendah sehingga perlu untuk melakukan glikolisis yang akan menghasilkan energi. AMP akan mempromosikan perubahan konfigurasi enzim PFK dari T state menuju R state.
Pyruvat kinase
Piruvat kinase diaktivasi oleh F-1,6-BP pada hati. ATP dan Alanin (produk biosintetik dari piruvat) bertindak sebagai inhibitor alosterik pada piruvat kinase. Asetil koenzim A juga berperan dalam menginhibisi kerja dari pyruvat kinase. Aktivasi dari pyruvat kinase juga diinhibisi oleh aktivitas cAMP protein kinase melalui kerja hormon glukagon (Bryant Miles,2003)
Ringkasan dan kesimpulan
Proses glikolisis
Enzim kunci yang berperan dalam proses glikolisis adalah:
a. Heksokinase
Enzim heksokinase berperan dalam mengkatalisis reaksi posforilase pertama, yaitu mengubah glukosa menjadi glukosa-6-posfat:

Heksokinase memerlukan logam dengan ion divalent seperti Mg2+ untuk dapat teraktivasi. Ion logam ini dapat membentuk kompleks dengan ATP. Heksokinase diinhibisi oleh produk yang dikatalisis yaitu glukosa-6-phosphate. Konsentrasi yang tinggi dari glukosa-6-phosphate dapat memberikan signal bahwa sel tidak memerlukan lagi glukosa untuk dijadikan energi.
b.Phosphofructokinase (PFK)
Enzim PFK digunakan pada reaksi fosforelasi II yang mengubah fruktosa 6-fosfat menjadi fruktosa 1,6 bifosfat. Reaksi fosforelasi yang dikatalisis oleh fosfofruktokinase adalah reaksi yang memanfaatkan ATP sebagai sumber fosfat.
c. Pyruvat kinase
Piruvat kinase diaktivasi oleh F-1,6-BP pada hati. ATP dan Alanin (produk biosintetik dari piruvat) bertindak sebagai inhibitor alosterik pada piruvat kinase. Asetil koenzim A juga berperan dalam menginhibisi kerja dari pyruvat kinase. Aktivasi dari pyruvat kinase juga diinhibisi oleh aktivitas cAMP protein kinase melalui kerja hormon glukagon.
Proses glukoneogenesis
Enzim kunci yang berperan dalam proses glukoneogenesis adalah:
a.Glucose-6-phosphatase
Glukosa-6-posfatase bekerja mengubah glukosa-6-posfat menjadi glukosa .Glukosa-6-fosfatase merupakan enzim yang ditemukan banyak pada hati dan ginjal. G6Pase ini terikat pada membran yang terhubung dengan retikulum endoplasma. Penyusun dari G6Pase bergantung dari integritas membran mikrosom. Katalisis G6Pase merupakan reaksi pertukaran antara glukosa dan G6P dengan bukti, yaitu bahwa enzim ini membentuk fosfoenzim intermediate. Inhibitor Glc-6-P diharapkan menghambat G6Pase untuk utuh tetapi tidak mengganggu mikrosom.
b. fructose-1,6-bisphosphatase
Fructose-1,6-bisphosphatase berfungsi mengkatalisis reaksi pengubahan fruktosa-1,6-bifosfat menjadi fruktosa-6-posfat.
c. Inhibisi oleh fruktosa-2,6-bisfosfat
Fruktosa-2,6-bisfosfat merupakan inhibitor kompetitif. Proses inhibisi dapat terjadi karena fruktosa-1,6-bisfosfatase bersifat alosterik.
d.Phosphoenolpyruvat carboxykinase
Proses Glukoneogenesis melibatkan 4 jenis enzim kunci, salah satunya adalah phosphoenol pyruvat carboxykinase (PEPCK) yang mengkatalisis pembentukan phosphoenol piruvat dari oksaloasetat.
REFERENSI
Aschenbach WG, Suzuki Y, Breeden K, Prats C, Hirshman MF,Dufresne SD, Sakamoto K, Vilardo PG, Steele M, Kim JH,Jing SS, Goodyear LJ & DePaoli-Roach AA (2001) The muscle-specific protein phosphatase PP1G/R GL(GM) is essential for activation of glycogen synthase by exercise. Journal of Biological Chemistry 276, 39959–39967.
Bloch G, Chase JR, Meyer DB, Avison MJ, Shulman GI & Shulman RG (1994) In vivo regulation of rat muscle glycogen resynthesis after intense exercise. American Journal of Physiology 266, E85–E91.
Brady MJ & Saltiel AR (2001) The role of protein phosphatase-1 in insulin action. Recent Progress in Hormone Research 56,157–173.
Carling D & Hardie DG (1989) The substrate and sequence specificity of the AMP-activated protein kinase. Phosphorylation of glycogen synthase and phosphorylase kinase.Biochimica et Biophysica Acta 1012, 81–86.
Chasiotis D, Brandt R, Harris RC & Hultman E (1983a) Effects of beta-blockade on glycogen metabolism in human subjects during exercise. American Journal of Physiology 245, E166–E170.
Chasiotis D, Sahlin K & Hultman E (1982) Regulation of glycogenolysis in human muscle at rest and during exercise. Journal of Applied Physiology 53, 708–715.
Chasiotis D, Sahlin K & Hultman E (1983b) Regulation of glycogenolysis in human muscle in response to epinephrine infusion. Journal of Applied Physiology 54, 45–50.
Chen Z, Heierhorst J, Mann RJ, Mitchelhill KI, Michell BJ,Witters LA, Lynch GS, Kemp BE & Stapleton D (1999) Expression of the AMP-activated protein kinase beta1 and beta2 subunits in skeletal muscle. FEBS Letters 460, 343–348.
Cohen P (1978) The role of cyclic-AMP-dependent protein kinase in the regulation of glycogen metabolism in mammalian skeletal muscle. Current Topics in Cellular Regulation 14, 117–196.
Danforth WH (1965) Glycogen synthase activity in skeletal muscle. Interconversion of two forms and control of glycogen synthesis. Journal of Biological Chemistry 240, 588–593.
Day, C., Cauturight, T., Provost, J.and Bailey, C.J. 1990. Hypoglycemic effect of momordica charantia extracts. Planta Med. 56:426-429
Grégoire PE: Biochimie Pathologique. Presses Académiques Européennes, Brussels, 1970, p. 368
Hiraga A & Cohen P (1986) Phosphorylation of the glycogenbinding subunit of protein phosphatase-1G by cyclic-AMPdependent protein kinase promotes translocation of the phosphatase from glycogen to cytosol in rabbit skeletal muscle.European Journal of Biochemistry 161, 763–769.
Hojlund K, Staehr P, Hansen BF, Green KA, Hardie DG, Richter EA, Beck-Nielsen H & Wojtaszewski JFP (2003) Increased phosphorylation of skeletal muscle glycogen synthase at NH2- terminal sites during physiological hyperinsulinemia in type 2 diabetes. Diabetes 52, 1393.
Imparl- Radoservich J, Deas S, Polansky MM, Daedke DA, Ingebruten TS, Anderson RA, Graves DJ. Regulation of phosphotyrosine phosphate and insulin receptor kinase by factions and from cinnamon: implications for cinnamon regulation of insulin signaling hormone Res 50: 177-182-1998.
Jiao Y, Shashkin P & Katz A (2001) A new glycogen synthase activity ratio in skeletal muscle: effects of exercise and insulin. Life Sciences 69, 891–900.
Liemans V, Sener A, Malaisse WJ: New sensitive assay of fructose-1,6-diphosphatase activity. Med Sci Res 22: 259–261, 1994
Marchand-Brustel Y, Gautier N, Cormont M & Van Obberghen E (1995) Wortmannin inhibits the action of insulin but not that of okadaic acid in skeletal muscle: comparison with fat cells.Endocrinology 136, 3564–3570.
Markuns JF, Wojtaszewski JF & Goodyear LJ (1999) Insulin and exercise decrease glycogen synthase kinase-3 activity by different mechanisms in rat skeletal muscle. Journal of Biological Chemistry 274, 24896–24900.
Montell E, Arias A & Gomez-Foix AM (1999) Glycogen depletion rather than glucose 6-P increments controls early glycogen recovery in human cultured muscle. American Journal of Physiology 276, R1489–R1495.
Nielsen JN, Derave W, Kristiansen S, Ralston E, Ploug T & Richter EA (2001) Glycogen synthase localization and activity in rat skeletal muscle is strongly dependent on glycogen content. Journal of Physiology (London) 531, 757–769.
Parker GJ, Lund KC, Taylor RP & McClain DA (2003) Insulin resistance of glycogen synthase mediated by O-linked Nacetylglucosamine. Journal of Biological Chemistry 278,10022–10027.
Parker PJ, Caudwell FB & Cohen P (1983) Glycogen synthase from rabbit skeletal muscle; effect of insulin on the state of phosphorylation of the seven phosphoserine residues in vivo. European Journal of Biochemistry 130, 227–234.
Roach PJ (2002) Glycogen and its metabolism. Current Molecular Medicine 2, 101–120.
Roach PJ & Larner J (1976) Rabbit skeletal muscle glycogen synthase. II. Enzyme phosphorylation state and effector concentrations as interacting control parameters. Journal of Biological Chemistry 251, 1920–1925.
Roach PJ, Takeda Y & Larner J (1976) Rabbit skeletal muscle glycogen synthase. I. Relationship between phosphorylation state and kinetic properties. Journal of Biological Chemistry 251, 1913–1919.
Sakamoto K, Aschenbach WG, Hirshman MF & Goodyear LJ (2003) Akt signaling in skeletal muscle: Regulation by exercise and passive stretch. American Journal of Physiology 285, E1081–E1088.
Sakamoto K, Hirshman MF, Aschenbach WG & Goodyear LJ (2002) Contraction regulation of Akt in rat skeletal muscle. Journal of Biological Chemistry 277, 11910–11917.
Skurat AV & Dietrich AD (2003) DYRK Family protein kinases phosphorylates Ser-640 in muscle glycogen synthase. Diabetes 52, 1852.
Sener A, Malaisse WJ: Kinetics and specificity of human B-cell glucokinase: relevance to hexose-induced insulin release. Biochim Biophys Acta 1312: 73–78, 1996
Wojtaszewski JFP, Higaki Y, Hirshman MF, Michael MD,Dufresne SD, Kahn CR & Goodyear LJ (1999a) Exercise modulates postreceptor insulin signaling and glucose transport in muscle-specific insulin receptor knockout mice. Journal of Clinical Investigation 104, 1257–1264.
Wojtaszewski JFP, Jorgensen SB, Hellsten Y, Hardie DG & Richter EA (2002) Glycogen-dependent effects of 5-aminoimidazole- 4-carboxamide (AICA)-riboside on AMP-activated protein kinase and glycogen synthase activities in rat skeletal muscle. Diabetes 51, 284–292.
Wojtaszewski JFP, Lynge J, Jakobsen AB, Goodyear LJ & Richter EA (1999b) Differential regulation of MAP kinase by contraction and insulin in skeletal muscle: metabolic implications. American Journal of Physiology 277, E724–E732.
Wojtaszewski JFP, Nielsen P, Kiens B & Richter EA (2001) Regulation of glycogen synthase kinase-3 in human skeletal muscle: effects of food intake and bicycle
Sener A, Malaisse WJ: Hexose metabolism in pancreatic islets. Metabolic and secretory responses to D-fructose. Arch Biochem Biophys 261: 16–26, 1988
Malaisse WJ, Malaisse-Lagae F, Davies DR, Vandercammen E, Van Schaftingen E: Regulation of glucokinase by a fructose-1-phosphatesensitive protein in pancreatic islets. Eur J Biochem 190: 539–545,1990
Sener A, Malaisse WJ: Hexose metabolism in pancreatic islets: apparent dissociation between the secretory and metabolic effects of D-fructose. Biochem Mol Med 59: 182–186, 1996
Qin B, Nagasaki M, Ren M, et al. Cinnamon extract prevents the insulin resistance induced by high-fructose diet. Horm Metab Res 2004; 36:119-25.
Qin B., Nagasaki, V., 2003. Cinnamon extract oshida, Y., Sato Y., 2003.Cinnamon extract potentiates in vivo insulin regulated glucose utilization via enhancing insulin signaling in rats. Diabetes Research and clinical practice 62, 139-148
Komentar
Posting Komentar